ALTERING VOLATILE PRODUCTION IN OILSEED RAPE BY INTROGRESSION OF GENES FROM WILD BRASSICAS
Robert P Bradburne, Ruth Magrath and Richard F Mithen
Brassica and Oilseeds Department, John Innes Centre, Norwich, NR4 7UH, UK
ABSTRACT
In this study we have introgressed genes which alter aliphatic glucosinolate (GSL) production from a wild Brassica oleracea and a wild Brassica rapa plant into agronomically acceptable breeding lines of oilseed rape. The positions of these genes in the Brassica napus (oilseed rape) genome have been located through construction of a Restriction Fragment Length Polymorphism map of a Doubled Haploid plant population. We have also produced lines with altered GSL breakdown rates through introgression of genes involved in the myrosinase system. Analysis of plant lines differing in volatile production with respect to the interactions of these plants with generalist and specialist pests is discussed.
KEYWORDS Brassica napus glucosinolate myrosinase pests
INTRODUCTION
Glucosinolates (GSL) are thioglycosides found in plants throughout the order Capparales (Chew 1988). In most Capparalean families, indole and aromatic GSLs are prevalent, but the Brassicacae are distinct in producing a large amount of aliphatic GSLs in their seeds and vegetative tissues. These aliphatic GSLs are highly variable due to the large number of different side chain structures that they can possess. This in turn leads to a wide variety of biological functions of both the intact GSLs and also their breakdown products.
Oilseed Rape (Brassica napus) contains mostly 2-hydroxy-but-3-enyl and 2-hydroxy-pent-4-enyl GSLs in its seeds and leaves (Magrath et al. 1994). These both produce goitrogenic breakdown products upon ingestion, limiting the seed meal’s use in animal feed (Duncan 1992). To increase the value of seed meal there has been extensive breeding to reduce the total seed GSL levels in winter oilseed rape varieties. This has been largely achieved through the use of alleles from the Polish variety Bronowski which greatly reduce the total levels of seed GSLs (Campos and Mithen 1996). However, the intense use of this line in later breeding programs has meant that there is very little genetic variation in modern oilseed rape cultivars for GSL biosynthesis.
GSLs often act as defensive compounds to deter generalist herbivores from eating the plant tissues which contain them (Chew 1988). Studies on wild populations of Brassicas have shown that there can be selection for different GSL profiles under different herbivore pressures (Giamoustaris et al. 1995). Therefore one of the aims of this study has been to try to change the GSLs present in agronomically acceptable lines of winter oilseed rape to reduce feeding by generalist pests such as rabbits and woodpigeons. In doing this we have also introduced a novel GSL, not previously found in any oilseed rape cultivars, into these new lines, and the effects of this on pest interactions have been studied.
Due to the extremely restricted genepool available in oilseed rape cultivars, it has been necessary to look to wild Brassica plants for the genes needed to change GSL production. B.napus is an amphidiploid species, containing the equivalent of an entire genome from both Brassica oleracea and Brassica rapa (Parkin et al. 1995) and so representatives of these two species were chosen to form a resynthesised B.napus plant which could then be used in a breeding program to produce agronomically acceptable lines (summarised in Fig. 1).
By studying the lines produced at different stages in the breeding program, the genetics of aliphatic GSL biosynthesis in B.napus have been elucidated. The breakdown of these GSLs after tissue damage has also been studied. GSLs are broken down on tissue damage by the thioglucosidase “myrosinase”. They can form a range of breakdown products depending on hydrolysis conditions, but the main forms are isothiocyanates (ITC), thiocyanates, nitriles and epithionitriles (ETN) from non-hydroxylated forms and oxazolidine-2-thiones from hydroxy-glucosinolates (Chew 1988). These are highly biologically active compounds, and the consequences of these breakdown processes have also been analysed in this study.
METHODS
Production of novel oilseed rape lines.
Figure 1 shows the breeding program used in this study. (from Bradburne et al. 1999)
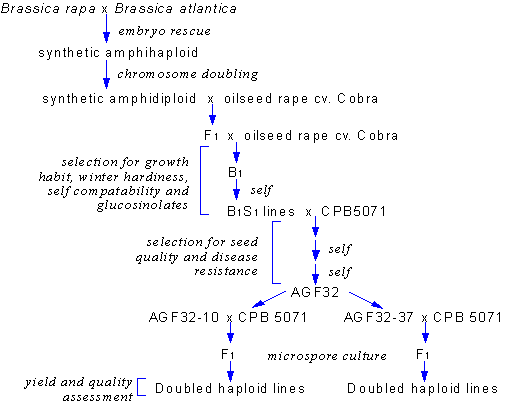
Figure 1. Breeding program used to produce novel oilseed rape lines.
Brassica atlantica is a form of B. oleracea which is found in Tunisia. The embryo rescue, colchicine doubling, and Doubled Haploid (DH) production techniques are described in Mithen and Magrath (1992) and Magrath et al. (1994). Throughout the breeding program the plants were analysed for GSL content by High Performance Liquid Chromatography (HPLC) as described in Magrath et al. 1993 and selected for those with high but-3-enyl and prop-2-enyl GSL levels in their leaves and low seed GSLs.
Analysis of Doubled Haploid lines.
Volatile production.
Total leaf GSL levels and breakdown rates were studied using the HPLC method outlined above, allowing 120 seconds of hydrolysis in water before the addition of the methanol in the breakdown studies. The volatiles were collected in the headspace over rehydrated, lyophilised leaf material using a Solid Phase Microextraction probe and analysed by Gas Chromatography-Mass Spectroscopy.
Genetic analysis.
DNA was exctracted from each of the 85 DH lines, restricted, blotted and probed with Restriction Fragment Length Polymorphism (RFLP) markers as described in Bradburne et al. (1999). An RFLP map was then constructed with reference to the maps of Sharpe et al. (1995) and Bohuon et al. (1996) and using the data from the GSL and volatile analysis, the loci responsible for altering volatile production were positioned. This was done using the Mapmaker and Mapmaker QTL packages (Lander et al. 1987)
Animal interaction studies.
Pigeon and rabbit damage was assessed in field plots on both the F32-10 parent and selected DH lines in 1996-7 and 1997-8 in comparison with the standard cultivar Apex.
Flea beetle damage was scored on the 1997-8 pigeon damage assessment plots in October 1997.
RESULTS
Altered glucosinolate and volatile production.

Figure 2. DH line f32-10 212 (b), showing a contrasting glucosinolate profile from the standard cultivar Apex (a) . Error bars are ± 1 S.E.
A subset of the DH lines exhibited a very different GSL profile from standard oilseed rape in their leaves (figure 2). The major changes are: i) a reduction in proportion of hydroxylated GSLs, ii) a shift in the major GSL class present from pentyl GSLs to butyl GSLs, iii) the introduction of prop-2-enyl GSL into the spectrum. All combinations of these changes were observed within the DH population.
The volatiles
produced by the same lines shown in figure 2 are shown in figure 3. 
Figure 3. The volatile compouds produced from leaves of oilseed rape cv. Apex (a) and DH line F32-10 212 (b). Error bars are ± 1 S.E. Note the different scales of (a) and (b).
The amount of volatiles produced by different DH lines was much more variable than the amount of GSLs produced. F32-10 212 produced 10 times the levels of volatiles that Apex produced, but only 1.5 times the amount of GSLs. Other lines produced fewer volatiles than would be expected from their GSL profiles (data not shown). Also different DH lines varied in the proportion of ITC:ETN produced (figure 3), although this was more variable within lines. Only very low levels of prop-2-enyl ITC were detected from any of the lines.
Genetics of glucosinolate and volatile production
On average about 24% of the genome of each DH line was derived from the wild Brassica spp., with an average of 31% wild alleles present on the linkage groups containing GSL biosynthetic loci. In all, four loci were found to be important in producing the phenotypes observed in the DH population, one allele for lower levels of hydroxylation having become fixed in the breeding scheme at the “AGF32” stage. Several DH lines exhibited yields as high as the parental lines in field trials, indicating that none of the GSL biosynthesis loci are linked to agronomically deleterious genetic loci (Bradburne et al. 1999).
At present, the differences in GSL breakdown and volatile production observed in these lines cannot be attributed to well defined genetic loci.
Animal interactions.
Pigeon damage has been extremely light during the years of this study, but there has been reduction in damage seen both on the parental line, F32-10 and three of the DH lines with GSL profiles similar to that of F32-10 212 (figure 2b). Conversely, there was an increase in the amount of flea beetle damage on these DH lines in October (unpublished data).
CONCLUSIONS
This study has shown that it is possible to alter the spectrum of volatile chemicals that is produced by oilseed rape without reducing the agronomic quality of the lines. It has also been shown that not only can the relative proportions of the chemicals be changed, but novel volatiles can also be introduced into the system by introgression of alleles from related wild species. The breakdown of the GSLs in these lines appears to be a complex system, probably involving several genetic loci, and possibly also some environmental inputs. Myrosinases occur in distinct forms in different locations within plant tissue and are also strongly affected by the presence of cofactors such as L-ascorbate (Chew 1988) and any of these factors may be playing a role in the differences seen in the DH population.
Altering the total amount of GSLs in oilseed rape leaves has been shown to have opposing effects on the generalist and specialist herbivore pests of the crop (Giamoustaris and Mithen 1995), but this study also shows that altering the spectrum of GSL-derived volatiles while maintaining total GSL levels can have similar effects.
ACKNOWLEDGEMENTS
The authors would like to thank the Biotechnology and Biological Sciences Research Council, CPB-Twyfords Ltd and the Home Grown Cereals Authority for funding this work.
REFERENCES
Bohuon E, Keith D, Parkin I, Sharpe A, Lydiate D (1996) Alignment of the conserved C genomes of Brassica oleracea and Brassica napus. Theor Appl Genet 93, 833-9.
Bradburne R, Majer D, Magrath R, Werner C, Lewis B, Mithen R (1999) Winter oilseed rape with high levels of resistance to Pyrenopeziza brassicae derived from wild Brassica species. Plant Pathology (in press).
Campos de Quiroz H, Mithen R (1996) Molecular markers for low-glucosinolate alleles in oilseed rape (Brassica napus L.) Molecular Breeding 2, 277-81.
Chew F (1988) Biological effescts of glucosinolates. in Biologically Active Natural Products: Potential use in agriculture (ed. H. Cutler) pp.155-81.
Duncan A (1992)Glucosinolates. in Toxic Substances in Crop Plants pp.126-47.
Lander E, Green P, Abrahamson J, Barlow A, Daly M, Lincoln S, Newburg L (1987) Mapmaker; an interactive computer package for constructiong primary genetic linkage maps of experimental and natural populations. Genomics 1, 174-81.
Giamoustaris A, Raybould A, Mithen R (1995) Divergent selection for secondary metabolites between wild populations of Brassica oleracea and its implications for plant-herbivore interactions. Heredity 75, 472-84.
Giamoustaris A, Mithen R (1995) The effest of modifying the glucosinolate content of leaves of oilseed rape (Brassica napus ssp. oleifera) on its interaction with specialist and generalist pests. Annuals of Allied Biology 126, 347-63.
Magrath R, Herron C, Giamoustaris A, Mithen R (1993) The inheritance of aliphatic glucosinolates in Brassica napus. Plant Breeding 111, 55-72.
Magrath R, Bano F, Morgner M, Parkin I, Sharpe A, Lister C, Dean C, Turner J, Lydiate D, Mithen R (1994) Genetics of aliphatic glucosinolates. I. Side chain elongation in Brassica napus and Arabidopsis thaliana. Heredity 72, 290-9.
Mithen R, Magrath R (1992) Glucosinolates and resistance to Leptosphaeria maculans in wild and clutivated Brassica species. Plant Breeding 108, 60-8.
Parkin I, Sharpe A, Keith D, Lydiate D (1995) Identification of the A and C genomes of amphidiploid Brassica napus (oilseed rape). Genome 38, 1122-31.
Sharpe A, Parkin I, Keith D, Lydiate D (1995) Frequent nonreciprocal translocations in the amphidiploid genome of oilseed rape (Brassica napus). Genome 38, 1112-21.