SIMULATING GROWTH AND DEVELOPMENT OF CANOLA IN AUSTRALIA
1M. J. Robertson, 2J.F. Holland, 3J. A. Kirkegaard, and 4C. J. Smith
1Agricultural Production Systems Research Unit, CSIRO Tropical Agriculture, 206 Carmody Rd, St. Lucia, Queensland 4067, Australia
2NSW Agriculture, RMB 944, Tamworth, New South Wales 2340, Australia
3CSIRO Plant Industry, GPO Box 1600, Canberra, ACT 2601, Australia
4CSIRO Land and Water, GPO Box 1666, Canberra, ACT 2601, Australia
ABSTRACT
This paper describes the development and testing of a canola module for the Agricultural Production Systems Simulator (APSIM), which simulates crop development and growth of canola. Model parameters describing phenology, leaf area development, biomass production and partitioning, demand, uptake and retranslocation of nitrogen, and root growth and water extraction, were assembled from the literature and growth analysis experiments conducted in Australia. The model was tested against independent datasets from Australia (26 to 36o latitude), which varied in terms of nitrogen supply, water supply, sowing date, and variety. Grain yields, ranging from 30 to 500 gm-2, were simulated with a root man squared deviation of 45 gm-2 (15% of the observed mean). We discuss features of the model that require further development.
KEYWORDS: APSIM, phenology, canopy development, biomass accumulation, grain-filling
INTRODUCTION
There is an increasing recognition in agricultural research that a systems approach is needed in the search for strategies for more efficient production, improved risk management, and sustainable production systems. Dynamic simulation models can increase research efficiency through the analysis of system performance at different locations, and for varying season lengths, where climate is unpredictable and relatively variable. They can also assist in the interpretation of field experimentation dealing with crop responses to soil, management, genetic or environmental factors. Canola production is similar to other cropping systems in that alternative management strategies must consider production, economic and environmental dimensions.
The cropping systems model, APSIM (Agricultural Production Systems Simulator), allows models of crop and pasture production, residue decomposition, soil water and nutrient flow, and erosion to be configured to simulate various production systems (McCown et al., 1996) including crop sequences and intercropping, and soil and crop management, to be dynamically simulated using conditional rules. APSIM was developed in response to previous efforts on modelling crop production, which had centred on the use of individual crop models responding to climate, water, nitrogen and management. APSIM is able deal with other features of the cropping system such as crop sequences, intercropping, crop residue management, and feedback between the crop and soil fertility status. This paper describes the development and validation of APSIM-Canola, a plug-in/pull-out module of APSIM, which simulates crop development and growth of canola.
Models of rapeseed production have been developed by Husson (1998) and Gabrielle et al. (1998a,b), using the CERES framework. Both models simulate biomass and nitrogen dynamics in response to climate, nitrogen and water supply, however they are limited in that they concentrate on winter types of rapeseed, and have not been tested on severely drought-stressed crops that are common in dryland environments in Australia.
The purpose of this paper is to describe the functional basis of the canola module, and report its ability to predict crop growth, development, yield and nitrogen uptake for spring cultivars in Australia under dryland and irrigated production systems. Demonstration of the application of the model to issues of canola production in the farming system can be found in companion papers at this conference by Holland et al. (1999) and Robertson et al. (1999).
MODEL DESCRIPTION
Table 1: Key input parameters and sources for the canola module. Those parameters derived by calibration using datasets P1 and P2 in Table 2 are indicated.
|
Parameter description |
Value |
Source |
|
Phenology |
|
|
|
Base, optimum and maximum temperature |
0, 25, 35 oC |
Virgil et al. 1997 |
|
Leaf Area Development |
|
|
|
Rate of node appearance on the main stem |
75 oCd pre stem elong, 20 oCd per node post stem elongation |
Morrison and McVetty (1991); Nanda et al. (1995) |
|
Number of leaves per plant per main stem node |
2.0 leaves per node |
Calibration |
|
Size of leaves (varies with node number) |
Range 800-50000 mm2 |
Nanda et al. (1995), P1 |
|
Rate of leaf death from nodes on the main stem |
38 oCd |
Triboi-Blondel (1988) |
|
Maximum specific leaf area for new leaf (varies with LAI) |
16000 to leaf 14 then declining |
Gabrielle et al (1998b) eqn 7 |
|
Biomass Accumulation |
|
|
|
Radiation use efficiency |
1.2 g MJ-1 (10% lower during pod-fill due to pod interception) |
Morrison and Stewart (1995); Mendham et al. (1981); Wright et al. (1988) |
|
Extinction coefficient for green leaf |
0.60 |
Gabrielle et al. (1998b); Mendham et al. (1981) |
|
Extinction coefficient for pod |
0.35 |
Gabrielle et al. (1998b) |
|
|
|
|
|
Transpiration efficiency coefficient |
0.005 kPa |
Hocking et al. (1997a) |
Biomass Partitioning |
|
|
|
Fraction of DM allocated to leaves pre flowering |
0.7 |
Gabrielle et al. (1998b), P1 |
|
Rate of harvest index increase during pod fill |
0.01 d-1 |
P1, P2 |
|
Multiplier of grain weight to account for seed energy content |
1.5 |
Hocking et al. (1997a) |
|
Root:shoot ratio (varies with stage of growth) |
0.3 – 0.05 |
JA Kirkegaard (unpubl.) |
Water Uptake |
|
|
|
Root extraction front velocity |
10 mm d-1 |
Gabrielle et al. (1998b), JA Kirkegaard (unpubl.) |
|
Nitrogen Demand |
|
|
|
Tissue nitrogen concentration below which plant becomes stressed (varies with plant part and stage of growth) |
Range 7.5 – 1 % |
Hocking (1993) |
|
Minimum tissue nitrogen concentration (varies with plant part and stage of growth) |
Range 5.5 – 0.3 % |
Hocking (1997b) |
All crop modules in APSIM use a similar framework. They simulate crop development, growth, yield and nitrogen accumulation (including fixation for legumes) in response to temperature, photoperiod, soil water and nitrogen supply, using a daily time-step. Daily maximum and minimum temperature, solar radiation and rainfall are climatic inputs. Crop phenology is divided into phases, the duration of each based on daily temperature and photoperiod. The duration from sowing to flowering is simulated as four phases: sowing to emergence; emergence to the end of the basic vegetative (or juvenile) period, which is a photoperiod-insensitive phase, and also dependent upon vernalisation in canola; a photoperiod-induced stage which depends upon the cultivar’s photoperiod sensitivity and which ends at floral initiation, and a floral development period which ends at flowering. The duration of grain-filling is determined by temperature. It is not the purpose of this paper to describe the phenology components in detail, as this is the subject of on-going research.
Leaf area development is described using functions for the appearance, expansion and senescence of leaves. More than one leaf may appear per node if assimilate supply permits, i.e. branching can occur. Leaf senescence is a function of age, light competition, drought and frost and has resultant impacts on leaf area, dry matter and plant nitrogen. Potential above-ground biomass production is predicted from leaf area index (and pod area index during grain filling), a radiation extinction coefficient and the crop’s radiation use efficiency. Actual daily biomass increase is calculated from the minimum of two potential crop growth rates, one determined by the intercepted radiation, limited by temperature and nitrogen stresses, and the other by soil water supply. This group of functions also partitions and retranslocates carbon between leaf, stem and grain.
The demand, uptake and retranslocation of nitrogen is also simulated, as is whole plant death due to stress. The crop has a defined minimum, critical and maximum N concentration for each plant part. Demand for nitrogen in each part attempts to maintain nitrogen at the critical (non stressed) level. Nitrogen demand on any day is the sum of the demands from the pre-existing biomass of each part required to reach critical N content, plus the N required to maintain critical N concentrations in that day’s potentially assimilated biomass. If nitrogen demand cannot be satisfied by mass flow to the roots from the soil, then it is supplied by active uptake.
The demand for assimilate for grain yield accumulation is defined by the linear increase with time of the harvest index. The ability to meet the grain yield demand is determined by the rate of biomass accumulation and the retranslocation of dry matter that had accumulated in the leaves and stems before the start of pod-filling. If assimilate supply is in excess of grain growth requirements, the excess is used for new leaf and stem growth. Parameter values for the key functional relationships that describe growth and development of the crop, listed in Table 1, were sourced from the literature (e.g. see the review by Mendham and Salisbury (1995)), and where unavailable, from unpublished experimental studies conducted by the authors.
Table 2: Details of data sets used to parameterise and test the canola module. Datasets P1 and P2 were used to derive model parameters, while datasets T1 to T7 are used for independent model testing.
|
Dataset |
Location |
Season |
Treatments |
Variety |
Reference |
|
P1 |
Moree, NSW (31.6oS, 119.2oE) |
1998 |
Dryland |
Hyola 42, Oscar |
JF Holland (unpubl.) |
|
P2 |
Gatton, Qld (27.6oS, 152.3oE) |
1998 |
Dryland |
Hyola 42, Oscar |
MJ Robertson (unpubl.) |
|
T1 |
Numurkah, Vic (36.1oS, 145.5oE) |
1985 |
Dryland, irrigated, 3 N rates |
Marnoo |
Wright et al. 1988, Smith et al. 1988 |
|
T2 |
Cootamundra, NSW (34.6oS, 148.0oE) |
1993-95 |
Dryland, previous crops |
Hyola, Oscar |
JA Kirkegaard (unpubl.) |
|
T3 |
Ginninderra, NSW (35.3oS, 149.1oE) |
1996-97 |
Dryland |
Dunkeld, Karoo, Oscar, Monty |
JA Kirkegaard (unpubl.) |
|
T4 |
Dalby, Qld (27.2oS, 151.3oE) |
1998 |
Dryland, 2 sowings |
Oscar, Hyola |
MJ Robertson (unpubl.) |
|
T5 |
Roma, Qld (26.6oS, 148.8oE) |
1998 |
Dryland |
Oscar, Hyola |
MJ Robertson and S. Cawley (unpubl.) |
|
T6 |
Tatura, Vic (36.4oS, 145.2oE) |
1987 |
Irrigated, 4 N rates |
Marnoo |
Taylor et al. 1991 |
|
T7 |
|
1989 |
Irrigated, 5 sowing dates |
Eureka |
Taylor and Smith 1992 |
MODEL TESTING
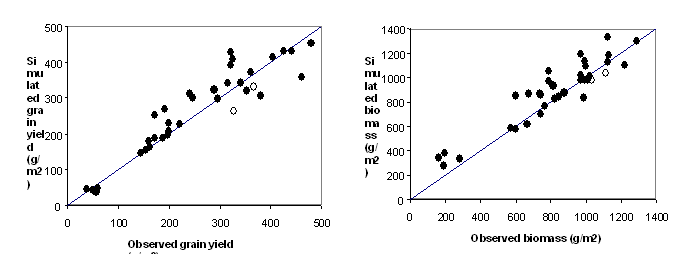
The 37 data points used to test the model ranged in grain yield from 38 to 480 g m-2, spanned latitudes from 36.4 to 26.6oS, and involved manipulations of agronomic factors like nitrogen supply, water supply, sowing date, and variety. Figs. 1a and b show that the model was able to credibly simulate the response to two important agronomic manipulations: sowing date and nitrogen, respectively. Over the 37 datasets, the model simulated yields and biomass at maturity over the range with no obvious bias (Fig. 2a,b). The root mean squared deviation of 45 and 134 g m-2 for grain yield and biomass, respectively, corresponds to 15% of the observed mean, and is of similar accuracy to other crop modules in APSIM. The scatter between observed and simulated biomass was higher than that for grain yield, partly due to uncertainties in the model in simulating variable leaf senescence and shedding late in grain filling.
CONCLUDING REMARKS
The present model appears to be capable of simulating variation in canola yields in response to location, sowing date, water supply, nitrogen supply, and cultivar in Australian environments, with reasonable accuracy. There are a number of features of the model that require further development and parameterisation. The seed oil concentration is known to vary with temperature, nitrogen supply and water stress, and such effects need to be accounted for by the model. While empirical relationships exist between the key variables and oil concentration (e.g Hocking et al. 1997a, Andersen et al. 1996, Gabrielle et al. 1997) there is no mechanistic approach available for simulation models. The parameters describing the phenological development, in relation to temperature, vernalisation and photoperiod, of the main Australian cultivars also need to be determined.
ACKNOWLEDGMENTS
Shayne Cawthray and Brett Cocks assisted with data collected at Gatton, Dalby and Roma. Thanks to Scott Cawley of the QDPI for permission to use data collected at Roma. This research was funded in part by the Grains Research and Development Corporation.
REFERENCES
Andersen, M.N., Heidmann, T., and Plauborg, F. 1996. The effects of drought and nitrogen on light interception, growth and yield of winter oilseed rape. Actu Agric. Scand. Sect. B. Soil and Plant Sci. 46:55-67.
Gabrielle, B., Denoroy, P., Gosse, G., Justes, E., and Andersen, M.N. 1998a. Development and evaluation of a CERES-type model for winter oilseed rape. Field Crops Res. 57:95-111.
Gabrielle, B., Denoroy, P., Gosse, G., Justes, E., and Andersen, M.N. 1998b. A model of leaf area development and senescence for winter oilseed rape. Field Crops Res. 57:209-22.
Hocking, P.J. 1993. Effects of sowing time and plant age on critical nitrogen concentrations in canola (Brassica napus L.). Plant and Soil 155/156:387-90.
Hocking, P.J., Kirkegaard, J.A., Angus, J.F., Gibson, A.H., and Koetz, E.A. 1997a. Comparison of canola, Indian mustard and Linola in two contrasting environments. I. Effects of nitrogen fertilizer on dry-matter production, seed yield and seed quality. Field Crop Res. 49:107-25.
Hocking, P.J., Randall, P.J., and DeMarco, D. 1997b. The response of dryland canola to nitrogen fertilizer: partitioning and mobilization of dry matter and nitrogen, and nitrogen effects on yield components. Field Crop Res. 54:201-20.
Holland, J. F., Robertson, M. J., Kirkegaard, J. A., Bambach, R., Cawley, S. 1999. Yield of canola relative to wheat and some reasons for variability in the relationship. These proceedings.
Husson, F., Wallach, D., and Vandeputte, B. 1998. Evaluation of CECOL, a model of winter rape (Brassica napus L.). European J. Agron. 8:205-14.
McCown, R.L.; Hammer, G.L.; Hargreaves, J.N.G.; Holzworth, D.P. and Freebairn, D.M. 1996. APSIM: A novel software system for model development, model testing, and simulation in agricultural systems research. Agric. Syst. 50: 255-271.
Mendham, N.J. and Salisbury, P.A. 1995. Physiology: Crop development, growth and yield. Pp. 11-64 in Brassica oilseeds: production and utilization, Editors D. S. Kimber and D. I. McGregor. Wallingford: CAB.
Mendham, N.J., Shipway, P.A., and Scott, R.K. 1981. The effect of delayed sowing and weather on growth, development and yield of winter oil-seed rape (Brassica napus). J. Agric. Sci., Camb. 96:389-416.
Morrison, M.J. and McVetty, P.B.E. 1991. Leaf appearance rate of summer rape. Can. J.Plant Sci. 71:405-12.
Morrison, M.J. and Stewart, D.W. 1995. Radiation-Use Efficiency in Summer Rape. Agron. J. 87:1139-42.
Nanda, R., Bhargava, S.C., and Rawson, H.M. 1995. Effect of sowing date on rates of leaf appearance, final leaf numbers and areas in Brassica campestris, B. juncea, B. napus and B. carinata. Field Crops Res. 42:125-34.
Robertson, M. J., Holland, J. F., Bambach, Cawthray, S. 1999. Response of canola and Indian mustard to sowing date in risky Australian environments: field and simulation analysis. These proceedings.
Smith, C.J., Wright, G.C., and Woodroofe, M.R. 1988. The effect of irrigation and nitrogen fertilizer on rapeseed (Brassica napus) production in south-eastern Australia. I. Nitrogen accumulation and oil yield. Irrig. Sci. 9:15-25.
Taylor, A.J, Smith, C.J, Wilson, I.B. 1991. Effect of irrigation and nitroegn fertilizer on yield, oil content, nitrogen accumulation and water use of canola (Brassica napus L.). Fertilizer Res. 29: 249-260.
Taylor, A. J. and Smith, C. J. 1992. Effect of sowing date and seeding rate on yield and yield components of irrigated canola (Brassica napus L.) grown on a red-brown earth in south-eastern Australia. Aust. J. Agric. Res. 43: 1629-41.
Triboi-Blondel, A. M. 1988. Mise en place et functionnement des feuilles de colza d’hiver: relations azote-carbone et senescence. Agronomie 8: 779-786.
Virgil, M.F., Anderson, R.L., and Beard, W.E. 1997. Base temperature and growing-degree-hour requirements for the emergence of canola. Crop Sci. 37:844-49.
Wright, G.C., Smith, C.J., and Woodroofe, M.R. 1988. The effect of irrigation and nitrogen fertilizer on rapeseed (Brassica napus) production in south-eastern Australia. I. Growth and seed yield. Irrig. Sci. 9:1-13.
Figure 1: (a) Simulation of response of grain yield to rate of nitrogen fertiliser applied under irrigated conditions at Tatura, Vic (dataset T6). Vertical bars represent twice the standard error of the mean. (b) Response to sowing date under irrigated conditions at Tatura, Vic (dataset T7).
Figure 2: Observed and predicted (a) grain yield (oven-dry) at maturity and, (b) total biomass at maturity for datasets P1 and P2 (hollow symbols) and T1 to T6 (filled symbols). The line is the 1:1 relationship.