ENZYMATIC TRANSFORMATION OF DESULFO-GLUCOSINOLATES IN PURE NITRILES USING A RECOMBINANT
b-O-GLUCOSIDASE
Renato Iori2 , Jean-Paul Wathelet1, Onofrio Leoni2, Nicolas Mabon1, Michel Marlier1 and Sandro Palmieri2
1 Faculté universitaire des Sciences agronomiques, Unité de chimie générale et organique
Passage des Déportés, n°2, B-5030 Gembloux, Belgium
e-mail: wathelet.jp@fsagx.ac.be;
2 Istituto Sperimentale per le Colture Industriali, Via di Corticella 133, Bologna, Italy
e-mail: istsci5@iperbole.bologna.it
ABSTRACT
The recombinant b-O-glucosidase (EC 3.2.1.21) from the thermophilic bacterium Tp8 does not catalyse glucosinolate degradation but transforms desulfo-glucosinolates viz. desulfo-sinigrin, desulfo-gluconapin, desulfo-progoitrin, desulfo-epiprogoitrin, desulfo-glucotropaeolin, desulfo-gluconasturtiin, desulfo-sinalbin, desulfo-limnantin, desulfo-glucoerucin and desulfo-glucoiberin in the corresponding pure nitriles (GC purity: +/- 99%). Prop-3-enyl, but-3-enyl, (2R)-2-hydroxybut-3-enyl, (2S)-2-hydroxybut-3-enyl, benzyl, phenethyl, 4-hydroxybenzyl, 3-methoxybenzyl, 4‑methylthiobutyl and 3-methylsulphinylpropyl cyanides have been respectively identified by GC‑MS. This thermostable enzyme is very different from myrosinase, a b-S-glucosidase (EC 3.2.3.1) present in Brassicaceae, which easily hydrolyses glucosinolates producing mainly a mixture of isothiocyanates, nitriles and eventually thiones. This endogenous b-S-glucosidase is totally inactive towards desulfo-glucosinolates, while the b-O-glucosidase tested is not active with synthetic S-glucose substrates such as the p-nitrophenyl-S-glucose. It is the first time that a b‑O‑glucosidase has been found to hydrolyse natural S-glucose substrates such as desulfo-glucosinolates. The possibility to produce pure nitriles by this way, especially chiral compounds ((2R)-2-hydroxybut-3-enyl, (2S)-2-hydroxybut-3-enyl...), appears to be interesting for application in fine chemistry. The effect of pH (2 to 9) and temperature (40 to 85°C) on the recombinant b‑O‑glucosidase activity has been determined with desulfo-sinigrin as substrate. Optimal activity of this thermostable enzyme is reached at pH 6.
KEYWORDS
Glucosinolate, myrosinase, sulfatase, glycosidase, Brassicaceae
INTRODUCTION
Glucosinolates (GLs) belong to a structurally homogeneous family of more than 100 miscellaneous secondary metabolites mainly contained in Brassicaceae (Fenwick et al., 1983). These plants even contain myrosinase (thioglucoside glucohydrolase, EC 3.2.3.1), which catalyses the hydrolysis of GLs to D-glucose, sulphate ion and a serie of sulphur and/or nitrogen containing compounds such as thiocyanates, isothiocyanates, oxazolidinethiones, nitriles, etc, depending on the substrate (GL) and reaction conditions. GLs are also substrates of a sulfatase (EC 3.1.6.1) extracted from Helix pomatia snail, producing desulfo-glucosinolates (DS-GLs) (Thies, 1979; Leoni et al., 1998). These desulfo-derivatives cannot be hydrolysed by myrosinase and other enzyme-catalysed reactions. Although it was reported that the recombinant b-O-glucosidase (EC 3.2.1.21), from the thermophilic bacterium Tp8 cloned into Escherichia coli, was not active with synthetic S-glucose substrates such as the p-nitrophenyl-S-glucose (Plant et al., 1988), we found that thioglucosidic bond of DS-GLs was hydrolysed by b-O-glucosidase-catalysed reaction.
MATERIALS and METHODS
Reagents
Recombinant b-O-glucosidase (Cod. G 6906) was purchased from Sigma-Aldrich. All chemicals were of analytical grade.
Glucosinolates and desulfo-glucosinolates
GLs were purified from natural sources (Table 1) according to the method reported by Visentin et al. (1992).
Table 1: Origin and structure of glucosinolates tested
|
Trivial name |
Structure of the side chain R |
Species of origin (ripe seeds) |
|
Sinigrin Gluconapin Progoitrin Epiprogoitrin Glucotropaeolin Gluconasturtiin Sinalbin Glucolimnantin Glucoerucin Glucoiberin |
CH2=CH-CH2- CH2=CH-CH2-CH2 (2R) CH2=CH-CHOH-CH2- (2S) CH2=CH-CHOH-CH2- Ph-CH2- Ph-CH2-CH2- 4-OH-Ph-CH2- 3-CH3O-Ph-CH2- CH3-S-(CH2)3-CH2- CH3-SO-CH2-CH2-CH2- |
Brassica juncea Brassica rapa cv. Silla Brassica napus cv. Jet Neuf Crambe abyssinica Lepidium sativum L. Nasturtium
officinalis Limnanthes alba Eruca sativa Iberis amara |
DS-GLs were produced by enzymatic catalysed reaction using Helix pomatia sulfatase (Type H1, Sigma Saint Louis, USA) following the procedure described by Leoni et al. (1998).
Activity assay
Enzymatic activity of b-O-glucosidase on DS-GLs was determined following the hydrolytic decomposition of pure desulfo-sinigrin by HPLC (column: RP 18 Lichrospher 125 x 4 mm, 5 µm; solvent: water/acetonitrile 98:2; flow: 1 ml).
Production of pure nitriles
Pure DS-GLs were transformed into nitriles by enzymatic reaction with the recombinant b-O-glucosidase. About 3 mg of DS-GLs were weighted in a 10 ml test tube and solubilized with 2 ml of a 50 mM phosphate buffer pH 6, then 100 µl of enzyme solution (0,9 mg/ml) were added. This mixture was heated in a water bath (55°C) for 15 hours. Nitriles were extracted with diethyl ether (V/V) and analysed for their identification.
Nitriles identification
Nitriles were identified by GC-MS (Hewlett-Packard HP 5890 gas chromatograph coupled with a HP 5989 A mass spectrometer). The operating conditions were as follows: HP-5 MS column (5% methylphenylsilicone, 28 m x 0.25 mm id x 0.1 µm); helium carrier gas (1.2 ml/min); splitless injector (275°C); oven temperature (12°C/min from 30°C to 120°C; 8°C/min from 120°C to 180°C; 12°C from 180°C to 290°C; 6 min at 290°C); electronic impact (70 eV); source (200°C); interface (275°C).
Optimal pH and temperature determination
The effect of pH (2 to 9) and temperature (40 to 85 °C) on the recombinant b-O-glucosidase activity have been determined with desulfo-sinigrin as substrate. In a test tube maintained at constant temperature were introduced 100 µl of desulfo-sinigrin (10 µmol/ml), 100 µl of glucosidase (0,9 mg/ml) and 1.8 ml of 50 mM phosphate buffer. The enzymatic activity was determined by HPLC as described above.
RESULTS and DISCUSSION
Production and identification of pure nitriles
Ten DS-GLs with different R groups (alcenyl, hydroxy-alcenyl, aromatic, thioalkyl and sulfonylalkyl) were transformed into nitriles by enzymatic reaction with the recombinant b-O-glucosidase (Figure 1).
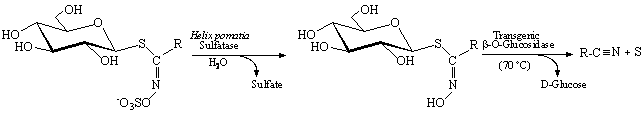 |
Figure 1: Transformation of glucosinolates into pure nitriles
Table 2 shows that all the compounds tested were hydrolysed and the correspondent nitriles (Chromatographic purity > 99%) have been produced. Finally, nitriles were characterised by GC-MS.
Table 2: Type and purity of produced nitriles
|
Name |
Nitrile identified |
Nitrile purity |
Isothiocyanate |
|
Desulfo-sinigrin Desulfo-gluconapin Desulfo-progoitrin Desulfo-epiprogoitrin Desulfo-glucotropaeolin Desulfo-gluconasturtiin Desulfo-sinalbin Desulfo-limnantin Desulfo-glucoerucin Desulfo-glucoiberin |
prop-3-enyl cyanide but-3-enyl cyanide (2R)-2-hydroxybut-3-enyl cyanide (2S)-2-hydroxybut-3-enyl cyanide benzyl cyanide phenethyl cyanide 4-hydroxybenzyl cyanide 3-methoxybenzyl cyanide 4-methylthiobutyl cyanide 3-methylsulfinylpropyl cyanide |
> 99% > 99% > 99% > 99% > 99% > 99% > 99% > 99% > 99% > 99% |
Not found Not found Not found Not found Not found Not found Not found Not found Not found Not found |
The physicochemical properties of recombinant thermostable b-O-glucosidase are very different from those of myrosinase, which is a b-S-glucosidase (EC 3.2.3.1) widespread in Brassicaceae. The latter can easily hydrolyse GLs producing mainly a mixture of isothiocyanates, nitriles and eventually thiones (Figure 2), whereas it is totally inactive towards DS-GLs. With this work we demonstrated for the first time that a recombinant b-O-glucosidase hydrolyses natural substrates such as DS-GLs which contain a b-thioglucosidic bond. The possibility to produce pure nitriles by this way, especially chiral compounds ((2R)-2-hydroxybut-3-enyl, (2S)-2-hydroxybut-3-enyl...), appears to be very interesting for fine chemistry applications and chromatographic standards production.
Figure 2: Enzymatic reactions with glucosinolates
 |
Optimal pH and temperature
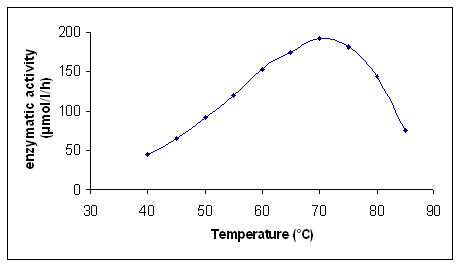
With the experiments reported in Figure 3 and 4, we have confirmed that the maximum enzyme activity is reached at pH 6 and the optimum enzymatic activity is at 70 °C. The latter finding confirmed that recombinant b-O-glucosidase is a thermoresistant enzyme as reported by Plant et al. (1988).
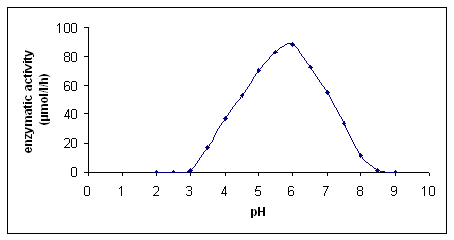 |
Figure 3: Influence of pH on the enzymatic activity
Figure 4: Influence of temperature on enzymatic activity
 |
CONCLUSIONS
In conclusion, we have demonstrated that nitriles can be obtained in pure form (chromatographic purity > 99%), without any formation of isothiocyanates or other volatile compounds from DS-GLs, using the recombinant b‑O‑glucosidase (EC 3.2.1.21) derived from the thermophilic bacterium Tp8. The production of pure and enantiopure nitriles, i.e. (2R)-2-hydroxybut-3-enyl cyanide and (2S)‑2‑hydroxybut-3-enyl cyanide, by this easy procedure appears to be very interesting for several fine chemistry applications, as well as for producing chromatographic standards. Finally, we want to emphasise that a new important catalytic function of commercial recombinant b-O-glucosidase in the hydrolysis of substrates containing thioglucosidic bond, such as DS-GLs, has been found.
REFERENCES
Fenwick G., Heaney R., Mullin W. (1983): Glucosinolates and their breakdown products in food and food plants. CRC, Critical Review in Food Science and Nutrition 18:123-201
Leoni O., Iori R., Haddoum T., Marlier M., Wathelet J-P, Rollin P. and Palmieri S. (1998): Approach to the use of immobilized sulfatase for analytical purposes and for the production of desulfo-glucosinolates. Industrial Crops and Products 7: 335-343
Plant A., Oliver J., Patchett M., Daniel R. and Morgan H. (1988): Stability and substrate specificity of a beta-glucosidase from the thermophilic bacterium Tp8 cloned into Escherichia coli. Archieves of Biochemistry and Biophysics 262 (1): 181-188
Thies W. (1979): Detection and utilisation of a glucosinolate sulfohydrolase in the edible snail Helix pomatia. Naturwissenschaften 66:364-365
Visentin M., Tava A., Iori R., Palmieri S., (1992): Isolation and identification of trans-4(methylthio)-3-butenyl glucosinolate from radish root (Raphanus sativus). Journal of Agricultural and Food Chemistry 40, 1687-1691