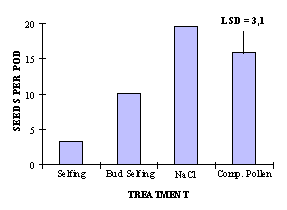
Fig. 1 : Seed production after selfing, bud selfing, application of NaCl solution and pollination with compatible pollen.
(F test 70,94***, p < 0,001)
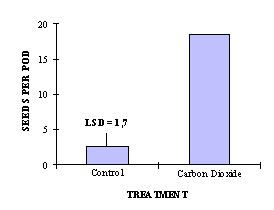
Fig. 2: Seed set after CO2 treatment.
(F test 353,18***, p < 0,001)
BREEDING HYBRID VARIETIES IN WINTER RAPESEED USING
RECESSIVE SELF-INCOMPATIBILITY
Silke Möhring, Elisabeth Esch, Günter Wricke
Abt. Angewandte Genetik, Universität Hannover, Herrenhäuser Str. 2, 30419 Hannover, Germany
e-mail: moehring@mbox.genetik.uni-hannover.de
ABSTRACT
For the efficient use of self-incompatibility in hybrid breeding selection has to be focused on inheritable, strong and stable expression of self-incompatibility to attain high hybridity levels. Different S alleles expressed in this way in a suitable genetic background would offer multiple possibilities to create single or three-way crosses as well as double cross hybrids.
Sodium chloride solution treatment of flowers seems to be an easy and appropriate method to overcome self-incompatibility for S line propagation.
Best hybrids showed yield performance 10 to 20 % higher than standard cultivars. Hybridity levels were influenced by environmental factors and genotype, nevertheless, in some combinations levels over 90 % have been achieved.
KEYWORDS
Brassica napus, self-incompatibility, sodium chloride, carbon dioxide, hybrids, hybridity
INTRODUCTION
Brassica oleracea and Brassica campestris, the ancestors of the allotetraploid Brassica napus have an efficient self-incompatibility system, whereas all cultivated forms of Brassica napus are self-fertile. Nevertheless self-incompatible genotypes could be found in rapeseed populations at a low level. Esch and Wricke (1995) reported a percentage of approximately 0,08. The advantages of S allele selection in breeding material are present good traits, in example low glucosinolate and erucic acid levels. Another possibility to gain new S alleles is their introduction in oilseed from the progenitor species.
Self-incompatibility can be used as pollination control mechanism in the production of hybrid rapeseed. Several methods are known to overcome self-incompatibility for the propagation of inbred lines. Monteiro et al. (1988) studied the effect of sodium chloride solution on self-incompatibility in Brassica campestris, Fu et al. (1992) and Esch (1994) carried out salt spraying experiments in Brassica napus. Nakanishi and Hinata (1975) and Palloix et al. (1985) found increased seed set in Brassica oleracea after exposure to carbon dioxide.
Significant heterosis for seed yield exists in oilseed rape. First experimental hybrids using self-incompatibility showed yield performance at least as good as the best cultivars (Esch and Wricke, 1995) or 20 % higher than the best parent (Banks and Beversdorf, 1994). These promising findings suggest further investigations towards the establishment of rapeseed hybrids based on self-incompatibility.
MATERIALS AND METHODS
Plant material and selection
Self-incompatible inbred lines with different S alleles were used in the experiments. Two different S alleles (Sx, Sy) derived from plant material described by Esch (1994). Additional S alleles (Sv, Sw) were selected from breeding material showing poor seed set after selfing in the field. Self-incompatibility was characterised by observing pollen tube growth by fluorescent microscopy (Kho and Baër, 1968) and self seed set.
Overcoming self-incompatibility
Sodium chloride solution treatment
Seed setting of a self-incompatible line was studied after selfing mature inflorescences, bud selfing, pollination with compatible pollen and selfing 1 h after stigma treatment with 8 % sodium chloride solution using a cotton swab (Monteiro et al., 1988). Each of these four treatments was carried out on 25 flowers of a lateral shoot of one plant. The experiment was repeated three times, each replication was represented by a single plant. The number of seeds per pod was used for the analysis of variance (ANOVA).
Carbon dioxide treatment
After the exposure of self-incompatible plants to carbon dioxide self seed set was determined. The experiment was carried out in two replications. Two plants per replication were tested, one plant was cultivated in normal atmosphere the other plant exposed to 6 % CO2 atmosphere and 80 % relative humidity after selfing. The CO2 atmosphere was built up in a growth chamber using dry ice where plants remained 24 h after selfing. Both treatments were carried out on 30 flowers manually selfed. Seed set per pod was evaluated in a one-factorial ANOVA.
Crosses in a factorial design
Five self-incompatible female inbreds of the fourth generation and five self-compatible male lines (L6 - L10) were crossed using a factorial mating design. Female lines SI 1, SI 8, SI 9, and SI 10 possessed the same S allele (Sx) while SI 11 carried the Sy allele. The hybrids were examined in field trials in two replications and three environments in 1998. Dry yield was subjected to a three-factorial ANOVA and compared to the average yield of four standard cultivars. The percentage of outcrossing (hybridity level) of several hybrids was determined at a random sample of 96 plants using the isozyme marker acid phosphatase.
RESULTS AND DISCUSSION
Selection
In addition to the S alleles Sx and Sy two new alleles (Sv, Sw) were found in breeding material to date. With diallel crosses it was confirmed that the four selected S alleles distinguish from each other. All S alleles showed recessive inheritance to self-compatibility. Hybrids derived from female lines carrying recessive S alleles and self-fertile male lines, fully regain self-compatibility. Seed setting of such hybrids after selfing is ensured even under bad weather conditions. In further studies additional S alleles should be selected to be used in different breeding strategies for the production of hybrid seed. The above mentioned alleles were introgressed into genetically diverse gene pools to produce improved self-incompatible lines. These lines with different genetic backgrounds can be used in most promising combinations in hybrid production. In order to facilitate detection of self-incompatible lines from backcross material an approach was made to find a molecular marker tightly linked to the S locus based on the PCR-RFLP method of Brace et al. (1993). This method works in Brassica napus but a codominant marker still has to be found.
Expression of self-incompatibility could be influenced by modifier genes (Hinata et al., 1983). Therefore a suitable genetic background should be found for each S alleles in which it will be strongly and stable expressed. A selection scheme enclosing selfing and fertility tests has to be developed to obtain such self-incompatible lines.
High temperature is able to weak the expression of self-incompatibility (Okasaki and Hinata, 1987). In one of our selection experiments self-incompatibility was influenced by different genotypes and temperatures. In another assay very different levels of self seed set were found within a progeny independent of temperature. In further studies the heritability of these different levels of pseudocompatibility will be examined.
Overcoming self-incompatibility
Sodium chloride solution treatment
The variation of seed set between the four distinct treatments was significant. A sodium chloride solution treatment is even superior to pollination with compatible pollen and also to bud selfing as can be seen in figure 1.
Propagation of self-incompatible lines in limited amounts for breeding purposes could be simply and fast realised with sodium chloride solution stigma treatment.
Spraying NaCl solution on open flowers of self-incompatible plants is an easier method but yielded in greenhouse experiments lower seed set than salt application to the stigma. The difficulty is to ensure that NaCl is applied directly to all stigmas by spraying. In our field trials salt treatment of an isolated self-incompatible line did not increase the self seed setting.
Fig. 1 : Seed production after selfing, bud selfing, application of NaCl solution and pollination with compatible pollen.
(F test 70,94***, p < 0,001)
Fig. 2: Seed set after CO2 treatment.
(F test 353,18***, p < 0,001)
Carbon dioxide treatment
An exposure of self-incompatible plants to a 6 % CO2 atmosphere lead to a high increase in seed set after selfing (Figure 2).
This method to overcome self-incompatibility is practically used in propagation of self-incompatible Brassica oleracea lines. High seed setting can be achieved after exposure to CO2. But this method needs a high technical effort whereas manual sodium chloride solution treatment is easily practicable and causes equivalent self seed set.
Crosses in a factorial design
The ANOVA of dry yield showed the presence of significant effects of males and females (p < 0,001) and a significant interaction (p < 0,001), indicating high general combining ability (gca) effects as well as specific combining ability (sca) effects. Therefore additive and dominant genetic effects influenced the yield.
These results differ from previous findings in yield performance of crosses in a factorial mating design (Möhring et al., 1997). In that experiment the yield was predominantly influenced by additive genetic effects. No significant dominance relationships among genes could be found. The absence of sca effects is rather uncommon for hybrids involving divergent parents. In 1998 the hybrids were grown in three environments. An explanation for the lack of sca effects might be that the experiment in 1997 was performed only in one environment. For that reason results of 1998 should be more representative.
Best hybrids resulted not always from crosses between parents showing high gca effects (Table 1). A parent with a relatively low gca was often involved in the best hybrid combinations (Table 2). According to existing sca effects the performance of hybrids can not be predicted on the basis of parental general combining ability alone. Several combinations have to be tested to find hybrids with high specific combining ability.
In opposition to the foregoing field trial in 1997 where best hybrids exceeded average standard cultivars by about 20 % (Möhring et al., 1997), best hybrids from 1998 showed a yield performance approximately 10 % higher than average standard cultivars (Table 2).
Tab. 1: Average of combination and gca effects for dry yield of parents.
Tab. 2: Dry yield and relative dry yield of best hybrids in comparison to standard cultivars.
|
Parent |
Dry yield average |
[ kg / plot ] gca effects |
|
SI 1 |
4,26 |
0,10 |
|
SI 8 |
4,31 |
0,15 |
|
SI 9 |
4,49 |
0,34 |
|
SI 10 |
4,44 |
0,28 |
|
|
|
|
|
Cultivar / Hybrid |
Dry yield [ kg / plot ] |
Rel. dry yield [ % ] |
|
Mohican |
4,13 |
95 |
|
Capitol |
4,10 |
94 |
|
Joker |
4,42 |
102 |
|
Life |
4,74 |
109 |
|
SI 11 |
3,28 |
- 0,87 |
|
L 6 |
3,91 |
- 0,24 |
|
L 7 |
4,34 |
0,18 |
|
L 8 |
4,32 |
0,17 |
|
L 9 |
4,15 |
- 0,01 |
|
L 10 |
4,06 |
- 0,10 |
|
SI 9 x L 7 |
4,73 |
109 |
|
SI 10 x L 9 |
4,78 |
110 |
|
SI 1 x L 8 |
4,75 |
109 |
|
SI 8 x L 7 |
4,70 |
108 |
Hybridity was also higher in 1997 with values about 90 to 99 %. Hybridity levels in 1998 reached a maximum about 70 to 80 %, perhaps due to bad weather conditions at the time of hybrid production. These moderate hybridity levels may have caused the lower yield performance in 1998.
Because of the requirement of strip planting of self-compatible pollinator rows alternating with self-incompatible female lines hybridity levels depend heavily on weather conditions. In three- or four-way crosses using different S alleles as proposed by Kott (1995) hybrids could be produced with blended seed for higher pollination efficiency. Seed amount of self-incompatible parental lines to produce such hybrids is much lower than for single cross hybrids. Therefore inbred line propagation could be easily performed with manual sodium chloride solution treatment in the greenhouse.
ACKNOWLEDGEMENT
The authors want to thank the Deutsche Saatveredelung GmbH, Lippstadt, and the German Bundesministerium für Bildung und Forschung for the support of this study.
REFERENCES
Banks, P. R. and W. D. Beversdorf 1994: Self-incompatibility as a pollination control mechanism for spring oilseed rape, Brassica napus L. Euphytica 75, 27-30.
Brace, J., D. J. Ockendon and G. J. King 1993: Development of a method for the identification of S alleles in Brassica oleracea based on digestion of PCR-amplified DNA with restriction endonucleases. Sexual Plant Reproduction 6, 133-138.
Esch, E. 1994: Untersuchungen zur Selbstinkompatibilität bei Brassica napus L. im Hinblick auf die Züchtung von Hybridsorten. Dissertation, Universität Hannover.
Esch, E. and G. Wricke 1995: Investigations on self-incompatibility in Brassica napus L. towards
hybrid breeding. Proceedings of the 9th International Rapeseed Congress, Cambridge, 83-85.
Fu, T.D., P. Si, X. Yang and G. Yang 1992: Overcoming self-incompatibility of Brassica napus by salt (NaCl) spray. Plant Breeding 109, 255-258.
Hinata, K., K. Okazaki and T. Nishio 1983: Gene analysis of self-compatibility in Brassica campestris var. yellow sarson (a case of recessive epistatic modifier). Proceedings of the 6th
International Rapeseed Conference, Paris, 354-359.
Kho, Y. O. and J. Baër 1968: Observing pollen tubes by means of fluorescence. Euphytica 17, 298-302.
Kott, L. S. 1995: Hybrid production systems based on self-incompatibility in oilseed Brassica. Proceedings of the 9th International Rapeseed Congress, Cambridge, 73-78.
Möhring, S., E. Esch, H. Busch and G. Wricke 1997: Selbstinkompatibilität in Winterraps (Brassica napus L.) als Bestäubungsmechanismus bei der Herstellung von Hybridsorten. 48. Tagung der Vereinigung östereichischer Pflanzenzüchter 1997, 77-80.
Monteiro, A. A., W.A. Gabelman and P.H. Williams 1988: Use of sodium chloride solution to overcome self-incompatibility in Brassica campestris. HortScience 23 (5), 876-877.
Nakanishi, T. and K. Hinata 1975: Self-seed production by CO2 gas treatment in self-incompatible cabbage. Euphytica 24, 117-120.
Okazaki, K. and K. Hinata 1987: Repressing the expression of self-incompatibility in crucifers by short-term high temperature treatment. Theoretical and Applied Genetics, 73, 496-500.
Palloix, A., Y. Herve, R.B. Knox and C. Dumas 1985: Effect of carbon dioxide and relative humidity on self-incompatibility in cauliflower, Brassica oleracea. Theoretical and Applied Genetics 70, 628-633.