CONDENSED TANNINS OF RAPESEED: NEW FINDINGS AND CHALLENGES
Marian Naczk1, Ryszard Amarowicz2, Fereidoon Shahidi3
1Department of Human Nutrition, St. Francis Xavier University, Antigonish, NS, Canada B2G 2W5
2Institute of Animal Reproduction and Food Research, Polish Academy of Sciences, Olsztyn, Poland
3Department of Biochemistry, Memorial University of Newfoundland, St. John’s, NF, Canada A1B 3X9
ABSTRACT
The utilization of rapeseed/canola as a source of food grade proteins is still limited due to the presence of glucosinolates, phytates, hulls and phenolics. Phenolic compounds contribute to the dark color, bitter taste and astringency of rapeseed protein products. The content of soluble condensed tannins in rapeseed /canola hulls is up to 15 times higher than the amounts reported previously. The hulls may also contain up to 5.5% insoluble condensed tannins. The crude tannin extract isolated from Cyclone canola hulls contained 47% tannins that were separated into four fractions on a Sephadex LH-20 column. The TLC studies revealed the presence of both oligomeric and more polymerized tannins. The chemical structure and contribution of these fractions to the taste of rapeseed protein products is still unknown. The titration curves (depicting the relationship between tannin content and the amount of tannin-protein complex formed at a standard protein concentration) were monitored using the protein precipitation assay and dye-labeled BSA assay. For both assays, we found the existence of a statistically significant (p# 0.001) correlation between the slope values and the tannin content in the hulls. We also found that the amounts of both precipitated tannins and precipitated dye-labeled BSA were higher at 40 0C than those precipitated at room temperature. This temperature sensitivity suggests that hydrophobic forces may play an important role in canola tannins-protein interactions.
KEYWORD: canola, tannin-protein interactions, oligomeric tannins, soluble tannins, insoluble tannins
INTRODUCTION
Tannins are complex polyphenolic compounds having molecular weights in the range 500 to 3000 daltons. They are widely distributed in foods and feeds of plant origin. The presence of condensed tannins was first reported by Bate-Smith & Ribereau-Gayon (1959). This finding was verified by Durkee (1971), who identified cyanidin, pelargonidin and an artifact n-butyl derivative of cyanidin in the hydrolytic products of rapeseed hulls. Later, Leung et al. (1979) reported that condensed tannins of rapeseed hulls contained leucocyanidin as their basic units. Advances in dehulling of rapeseed may soon bring about the introduction of dehulling to rapeseed/canola processing. The subsequent use of hulls as a component of feedstuff or food may be considered. Our studies (Naczk et al. 1994) has indicated that canola hulls contain up to 2% soluble tannins i.e. up to eight times more soluble tannins than that reported previously (Mitaru et al. 1982; Leung et al.. 1979).
MATERIALS AND METHODS
Hulls of Westar, Ebony, PR 3113 and Cyclone canola and Polo, Mar, Marita and Leo rapeseed varieties were prepared according to the procedure described by Sosulski and Zadernowski (1981). Hulls were extracted with hexane for 12 h using a Soxhlet apparatus and then dried at room temperature.
The soluble condensed tannins were extracted from hulls with 70% acetone as described by Naczk et al. (1994) and then assayed colorimetrically by the modified vanillin method (Price et al. 1978). The hull residue, after acetone extraction, was dried at room temperature and then finely ground. The soluble condensed tannins were also assayed by the proanthocyanidin method of Mole and Waterman (1987) as described by Naczk et al. (1994). The content of tannins were expressed in mg tannins/100g hulls.
The insoluble condensed tannins were assayed by the proanthocyanidin method of Mole and Waterman (1987) as follows. A 10 to 30 mg sample of finely ground, acetone extracted hulls, were extracted 6 times with a mixture of 1 mL of methanol and 10 mL of the 1-butanol-HCl reagent. The suspension was heated in sealed ampules for one hour in a boiling water bath and the contents were vortexed for 5 seconds every 10 minutes. The absorbance of the solution was measured at 550 nm against a reagent only blank. For A > 0.75 a dilution of the reaction mixture was made with 1-butanol. The content of insoluble tannins was expressed as mg tannins/100g hulls.
The canola tannins were purified and fractionated on a Sephadex LH-20 column according to the method of Strumeyer and Malin (1978). The separated fractions (in total) were examined using silica gel (Sigma) TLC methodology as described by Lea (1978).
The protein precipitating capacity of canola tannins was assayed by the protein precipitation method of Hagerman and Butler (1978) and by the dye-labeled BSA assay of Asquith and Butler (1985) with pH modifications as described by Naczk et al. (1996).
RESULTS AND DISCUSSION
The content of soluble condensed tannins in rapeseed/canola hulls ranged from 80 to up to 2500 mg /100g hulls as determined by the vanillin assay and ranged from 150 to up to 2000 mg/100g hulls as determined by the proanthocyanidin assay. These values are up to 15-times more than reported previously. These results suggest that both cultivar types and environmental growing conditions may affect the content of soluble tannins in hulls. The content of insoluble tannins ranged from 1500 to up to 5500 mg/100g hulls. The differences in the insoluble tannin content both between and within canola/rapeseed varieties are much lower than that found for soluble tannin contents.
Cyclone canola tannins were eluted into four fractions on a Sephadex LH-20 column using 50% acetone as eluent. Similar fractionation of condensed tannins, eluted from a Sephadex LH-20 column using 70% acetone, was reported by Czochanska et al. (1980). According to these authors tannins were fractionated according to their molecular size. The TLC chromatogram of each fraction of canola tannins revealed the presence of more polymerized tannins in Fractions I and II and the presence of oligomeric tannins in Fractions III and IV. Presence of catechins was not detected in these fractions.
Results of our studies indicates that the slope values of titration curves (depicting the amount of proteins or tannins precipitated as a complex depending on the concentration of canola tannins) were affected by both tannin and the protein used. A statistically significant correlation (p£ 0.001) existed between the slope values and tannin content in hulls. The slopes values of titration curves obtained using the precipitation assay ranged from 2.96 to 7.07, and those of titration curves obtained using dye-labeled BSA assay ranged from 35 to 200. The slope values of titration curves for tannins isolated from low-tannin hulls were higher than those found for tannins isolated from high-tannin hulls. The differences in the slope values of titration curves obtained for crude tannins isolated from low- and high-tannin hulls may be due to the existing differences in their affinities for proteins brought about by the differences in their molecular weights (Naczk et al., 1994). However, the chemical structure, polymer chain lengths, and molecular weights of canola/rapeseed tannins are still unknown.
Figure 1 shows the effect of temperature on the amount of tannin forming insoluble complex with BSA. We found that the amounts of precipitated tannins were higher at 40 oC than those precipitated at
room temperature (22 oC). A similar relationship was found using the dye-labeled assay, for the amount of proteins precipitated by tannins present. This temperature sensitivity suggests that hydro-
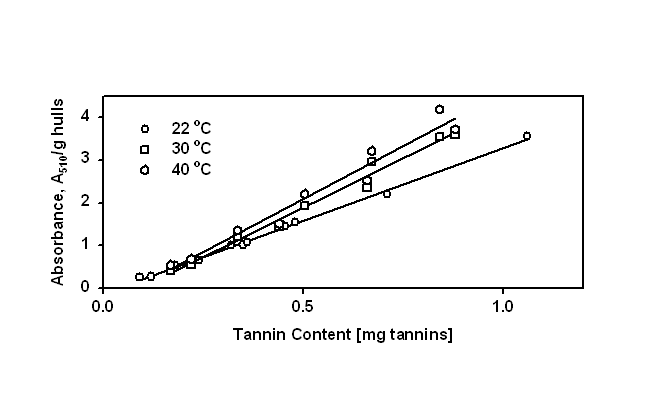
Figure 1. Effect of temperature on titration curves of a known amount of protein (1mg/mL)
with increasing amounts crude extracts of Cyclone canola tannins
phobic forces may play an important role in canola tannins-protein interactions.
ACKNOWLEDGEMENT
Marian Naczk thanks the Natural Sciences and Engineering Research Council of Canada (NSERC) for support in the form of a research grant.
REFERENCES
Asquith, T.N.; Butler, L.G. Journal of Chemical Ecology 1985, 11: 1535.
Bate-Smith, E.C.; Ribereau-Gayon, P. Qualitas et Plant Material Vegetabiles 1959, 5:189.
Czochanska, Z.; Foo, L.Y.; Newman, R.H.; Porter, L.J. Journal of Chemical Society Perkin Transactions 1980, 1: 2278,
Durkee, A. B. Phytochemistry 1971, 10: 1583.
Hagerman, A.E.; Butler, L.G. Journal of Agricultural and Food Chemistry 1978, 26:809.
Lea, A.G.M. Journal of the Science of Food and Agriculture 1978, 29:478.
Leung, J.; Fenton, T.W.; Mueller, M.M.; Clandinin, D.R. Journal of Food Science 1979, 44: 1313.
Mole, S.; Waterman, P.G. Oecologia 1987, 72:137.
Naczk, M.; Nichols, T.; Pink, D.; Sosulski, F. Journal of Agricultural and Food Chemistry 1994, 42:2196.
Naczk, M.; Oickle, D.; Pink, D.; Shahidi, F. Journal of Agricultural and Food Chemistry 1996, 44: 2144.
Price, M.L., van Scoyoc, S. and Butler, L.G. Journal of Agricultural and Food Chemistry 1978, 26:1214.
Strumeyer, D.H. and Malin, M.J. Journal of Agricultural and Food Chemistry 1978, 23: 1214.